

0 引言
混合发酵因可平衡系统营养成分、提升缓冲能力和调节微生物组成,是提高系统性能和运行稳定性的有效方法[1,2]。养殖废弃物和木质纤维素类原料是被广泛使用的发酵原料,其中养殖废弃物已成为主要的面源污染物,厌氧发酵是其主要的治理方式之一。多年生能源草本植物作为一类木质纤维素类原料,如杂交狼尾草(hybrid Pennisetum)、柳枝稷等,因其生物质量高和品质优良,成为一种潜在的沼气工程原料[3,4,5]。对于杂交狼尾草,目前已报道的年产生物质量最高为88 Mt/ha[6],产甲烷率为104 ~ 328 mL/g VS[3,7-9]。齐利格娃等[10]的研究表明,在中温厌氧干发酵工程中猪粪与稻草的最适挥发性固体(volatile solid, VS)配比为2:1,水力停留时间为36 d。在青贮草和牛粪混合发酵系统中适宜有机负荷为4.0 g VS/(L∙d),此时原料产甲烷率为349 mL/g VS[11]。XIE等[12]采用480 L 全混式反应器(continuously stirred tank reactors, CSTR)研究猪粪和青贮草混合发酵的稳定性,结果表明在VS比为1:1、有机负荷为1.74 g VS/(L∙d) 时系统可稳定运行。可见对于不同的混合发酵系统,因原料配比和运行条件变化,其运行性能存在差异。
微生物群落组成与原料配比和运行条件等因素息息相关,并影响系统运行性能。LI等[13]研究表明,在餐厨垃圾和纸张废弃物混合系统中,当纸张废弃物含量高于50%时,参与水解的微生物由碳水化合物/蛋白质水解菌属转向纤维素降解菌属,同时单糖在转化过程中直接降解为乙酸和H2,而不生成丁酸和丙酸。当果蔬废弃物和猪粪混合比例为70:30时,其优势细菌菌属为厚壁菌门(Firmicutes)和拟杆菌门(Bacteroidetes),产甲烷古菌属则是乙酸营养型和氢营养型产甲烷菌属共存;而当猪粪质量分数增加至50% ~ 60%时,系统产气量下降,与此对应的是氢营养型产甲烷菌属含量增加[14]。在玉米秸秆单独发酵系统中优势微生物为拟杆菌门(Bacteroidetes)和厚壁菌门(Firmicutes),而在玉米秸秆和牛粪混合比例为30:70时,系统的优势细菌更替为变形菌门(Proteobacteria)和螺旋体门(Spirochaetes)[15]。由此可见原料配比及有机负荷等因素都将影响厌氧发酵系统中微生物群落组成。基于此,本文采用生物测序技术,研究杂交狼尾草和牛粪混合发酵系统在不同原料配比和不同有机负荷下微生物群落演替规律,为通过调控混合发酵系统的运行条件改变微生物结构组成进而提高产气量提供理论支持。
1 实验部分
1.1 实验材料与接种物
Table 1 Characteristics of hybrid Pennisetum and cow manure表1 杂交狼尾草和牛粪的理化性质 |
| 原料 | 总固体质量分数TS / % | 挥发性固体质量 分数VS / % | 碳含量C / % | 氮含量N / % | 碳氮比C/N |
|---|---|---|---|---|---|
| 杂交狼尾草 | 15.87 ± 0.62 | 12.81 ± 0.45 | 39.59 ± 0.04 | 0.92 ± 0.02 | 43.28 ± 0.96 |
| 20.94 ± 0.84 | 18.17 ± 0.81 | 43.41 ± 0.54 | 0.70 ± 0.0 | 62.50 ± 2.68 | |
| 19.05 ± 1.15 | 16.48 ± 1.18 | 40.51 ± 0.33 | 0.66 ± 0.02 | 61.88 ± 2.51 | |
| 16.68 ± 0.49 | 13.61 ± 0.96 | 38.98 ± 0.57 | 0.74 ± 0.02 | 53.06 ± 2.31 | |
| 牛粪 | 14.42 ± 0.12 | 12.70 ± 0.01 | 41.32 ± 0.02 | 1.89 ± 0.04 | 21.87 ± 0.48 |
| 16.56 ± 0.31 | 14.46 ± 0.31 | 42.89 ± 0.28 | 2.16 ± 0.04 | 19.86 ± 0.52 |
接种物取自实验室的全混式反应器,用纤维素粉和蛋白胨长期驯化,接种物总固体(total solid, TS)及VS的质量分数分别为 (1.87 ± 0.03)%和 (1.10 ± 0.01)%。
1.2 实验装置与方法
半连续实验采用CSTRs[碧普(瑞典)有限公司],反应器有效容积为2 L,发酵温度为35 ± 1℃,定时搅拌(每隔3 min搅拌1 min)。产气量通过数据采集装置记录并保存,具体实验装置见图1。每个反应器中加入1 800 mL接种物,杂交狼尾草和牛粪的混合VS质量分数设定为1:1(GC11)和3:1(GC31),反应器的初始有机负荷(organic loading rate, OLR)为2.0 g VS/(L∙d),待系统稳定运行后,以0.5 g VS/(L∙d)为梯度,逐渐将其增加到5.5 g VS/(L∙d)。在OLRs为2.0 ~ 4.5 g VS/(L∙d) 时水力停留时间设为30 d,而当OLRs为5.0 g VS/(L∙d)和5.5 g VS/(L∙d) 时,进入系统的原料质量平均值分别为68.48 g/d和75.26 g/d,故该阶段水力停留时间分别为26 d和24 d。整个实验持续237 d。

Fig. 1 Laboratory set-up for semi-continuous experiments of anaerobic digestion: (a) whole reaction system; (b) 2 000 mL reactor图1 半连续进出料厌氧发酵实验装置:(a)整体反应系统;(b)2 000 mL反应器示意图 |
1.3 微生物群落组成分析方法
1.3.1 样品采集
杂交狼尾草与牛粪混合发酵系统分别在有机负荷为2.0 g VS/(L∙d)、2.5 g VS/(L∙d)、3.0 g VS/(L∙d)、3.5 g VS/(L∙d)、4.0 g VS/(L∙d)、4.5 g VS/(L∙d)、5.0 g VS/(L∙d)和5.5 g VS/(L∙d)时取样分析微生物组成,即于发酵进行的第21 d、63 d、92 d、119 d、144 d、173 d、206 d和228 d(GC11)/213 d(GC31)取样。
1.3.2 样品预处理及DNA提取
收集后的样品经离心弃去上清液,剩余固体用于微生物脱氧核糖核酸(deoxyribonucleic acid, DNA)提取。采用E.Z.N.ATM Mag-Bind Soil DNA Kit试剂盒(OMEGA)进行提取,并用琼脂糖凝胶电泳检测DNA完整性。
1.3.3 PCR扩增
聚合酶链式反应(polymerase chain reaction, PCR)体系如下:2×Taq master Mix,15 μL;Bar-PCR primer F(10 μmol/L),1 μL;Primer R(10 μmol/L),1 μL;Genomic DNA,10 ~ 20 ng;加水至30 μL。
PCR扩增反应条件如下:94℃预变性3 min;94℃变性30 s、45℃退火20 s、65℃延伸10 s,进行5个循环;94℃变性20 s、55℃退火20 s、72℃延伸30 s,进行20个循环;最后72℃延伸5 min,4℃保存。
细菌用V3-V4通用引物扩增:341F引物为CCTACGGGNGGCWG CAG;805R引物为GACT- ACHVGGGTATCTAATCC。
古菌用两轮槽式PCR扩增。第一轮是340F: CCCTAYGGGGYGCASCAG;1000R:GGCCATGC- ACYWCYTCTC。第二轮是349F:GYGCASCAGKC- GMGAAW;806R:GGACTACVSGGGTATCTAAT。
1.3.4 高通量测序分析及数据处理
高通量测序委托生工生物工程(上海)股份有限公司完成,采用Illumina Miseq 2X300bp平台测序,并完成数据处理。
数据库是16S细菌古菌核糖体数据库,包括RDP数据库(http://rdp.cme.msu.edu/misc/resources.jsp)、Silva数据库(http://www.arb-silva.de/)和NCBI 16S数据库(http://ncbi.nlm.nih.gov/)。
多样性指数分析采用mothur软件。群落分布丰度(community richness)为Chao1指数(http://www.mothur.org/wiki/Chao);群落分布多样性(community diversity)的指数有香农指数(Shannon)(http://www.mothur.org/wiki/Shannon)和覆盖度(coverage)(http://www.mothur.org/wiki/Coverage)。
2 结果与讨论
2.1 杂交狼尾草和牛粪混合发酵系统的微生物组成
前期研究结果表明,在杂交狼尾草和牛粪混合发酵系统中,当杂交狼尾草和牛粪混合VS比为1:1(GC11),将系统有机负荷从2.0 VS/(L∙d) 逐步升高至5.0 g VS/(L∙d)时发现,原料产气率和产甲烷率从526.33 mL/g VS和283.93 mL/g VS降至369.14 mL/g VS和210.64 mL/g VS,池容产气率则逐步升高,同时pH值、中间碱度/部分碱度(IA/PA)和挥发性脂肪酸/总碱度(VFAs/TA)等参数说明系统在该有机负荷范围内处于稳定运行状态;随着GC11系统继续增加有机负荷至5.5 g VS/(L∙d),系统池容产气率、原料产气率和产甲烷率降低,同时pH值降低,而VFAs和TAN浓度升高至3 509 ~ 4 427 mg/L和1 550 ~ 2 050 mg/L,IA/PA和VFAs/TA比值也表明系统处于不稳定状态,系统运行性能下降[16]。而当杂交狼尾草和牛粪混合VS比为3:1(GC31)时,系统可在有机负荷为2.0 ~ 4.5 g VS/(L∙d)时稳定运行。但前期研究缺乏对不同有机负荷和不同原料配比下发酵系统内微生物组成变化的分析,因此在前期研究的基础上对混合系统在不同有机负荷和原料配比下的微生物群落演替规律进行分析。
杂交狼尾草和牛粪混合发酵系统中,在不同混合配比和有机负荷条件下,门水平上的优势细菌群落和目水平上的优势古菌群落组成相同。在细菌门水平上,拟杆菌门(Bacteroidetes)、变形杆菌门(Proteobacteria)和厚壁菌门(Firmicutes)均为优势细菌,其相对丰度分别为24.98% ~ 59.84%、8.27% ~ 42.70%和12.92% ~ 31.58%(图2)。此外还有绿弯菌门(Chloroflexi)、互养菌门(Synergistetes)、疣微菌门(Verrucomicrobia)和浮霉菌门(Planctomycetes),其相对丰度分别为1.53% ~ 11.09%、0.95% ~ 5.28%、0.53% ~ 4.70%和0.21% ~ 1.93%。而在古菌目水平上,优势古菌有甲烷微菌目(Methanomicrobiales)、甲烷八叠球菌目(Methanosarcinales)、甲烷杆菌目(Methanobacteriales)、脱硫球菌目(Desulfurococcales)和Methanomassiliicoccales,总相对丰度为90.38% ~ 99.91%(图2)。
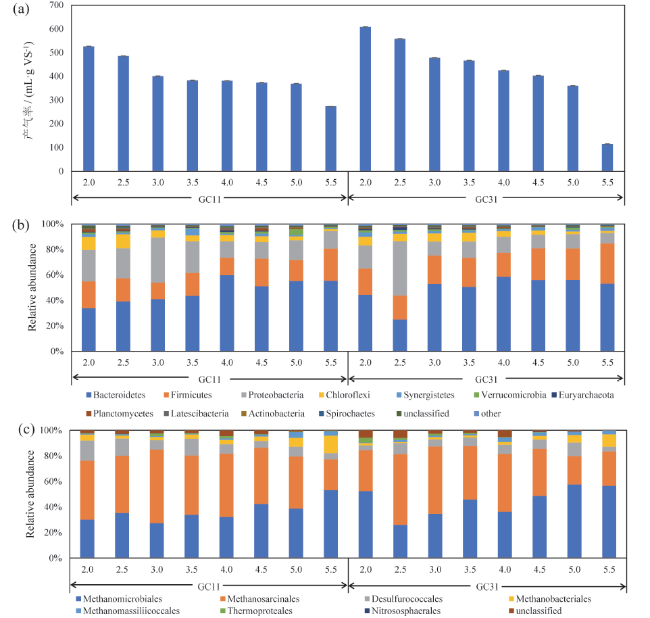
Fig. 2 The biogas yield (a), the relative abundance of bacteria in phylum level (b) and archaeal in order level (c) for co-digestion system图2 杂交狼尾草和牛粪混合发酵系统中的产气率(a)、细菌门水平上(b)和古菌目水平上(c)的微生物群落组成 |
2.2 杂交狼尾草和牛粪混合发酵系统中细菌演替规律
在杂交狼尾草和牛粪混合发酵系统中,属水平上细菌群落在不同有机负荷和原料配比下的变化情况如图3所示。在GC11系统中,梭菌属(Clostridum XIVa)在有机负荷低于5.0 g VS/(L∙d) 时相对丰度分别为0.04% ~ 0.59%,当有机负荷提高至5.5 g VS/(L·d)时,则升高至1.22%;普雷沃菌属(Prevotella)的相对丰度随着系统运行逐渐升高,当有机负荷低于5.0 g VS/(L∙d) 时该菌属相对丰度为0.04% ~ 1.81%,继续提高有机负荷至5.5 g VS/(L∙d),发现其相对丰度升高至3.66%,增幅达102.21%。在GC31系统中,梭菌属在有机负荷低于4.5 g VS/(L•d) 时,其相对丰度为0.10% ~ 0.48%,在5.0 ~ 5.5 g VS/(L·d) 时显著增加至4.16% ~ 4.82%,增幅达767% ~ 904%;而普雷沃菌属相对丰度在系统有机负荷低于4.5 g VS/(L·d) 时仅为0.01% ~ 0.09%,在5.0 ~ 5.5 g VS/(L•d) 时升高至4.44% ~ 4.93%。梭菌属Clostridum XIVa和 IV,是参与碳水化合物发酵产酸的菌属,可利用碳水化合物产生乙酸、丁酸和H2,是主要的产丁酸菌[17],通过丁酰辅酶A:醋酸辅酶A转移酶(butyryl CoA: acetate CoA transferase)途径代谢产丁酸。普雷沃菌属可在pH值4.6 ~ 5.0范围内生长,与纤维素水解菌协同作用实现植物细胞降解,丙酸是其主要发酵产物[18,19],同时该菌属与甲烷短杆菌属(Methanobrevibacter)具有很强关联性[19],从图4可见甲烷短杆菌属的相对丰度随之增加,增幅可达268%。可见,在GC11和GC31系统中贡献丙酸和丁酸累积的细菌菌属在有机负荷低于5.0 g VS/(L∙d)(GC11)或4.5 g VS/(L∙d)(GC31)时相对丰度较低,当有机负荷升高至5.5 g VS/(L·d)(GC11)或5.0 g VS/(L∙d)(GC31)时发生显著性增加,这与GC11和GC31系统分别在有机负荷为5.5 g VS/(L∙d) 和5.0 g VS/(L∙d) 时VFsA浓度升高结果相对应。
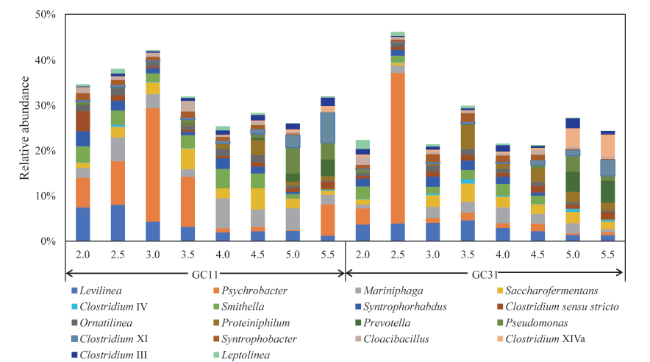
Fig. 3 Relative abundance of bacteria 16S rDNA gene in genus level for co-dogestion system图3 属水平上杂交狼尾草和牛粪混合发酵系统中细菌群落组成 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
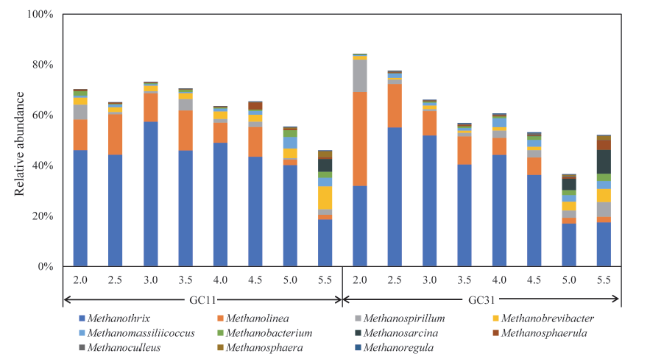
Fig. 4 Relative abundance of archaeal 16S rDNA gene in genus level for co-digestion system图4 属水平上杂交狼尾草和牛粪混合发酵系统中古菌群落组成 |
互养棍状菌属(Syntrophorhabdus)和互营杆菌属(Syntrophobacter)是互营丙酸氧化菌,与产甲烷菌协同降解丙酸。互营杆菌属含有4个种,均为中温互营丙酸降解菌,当存在硫酸盐和延胡索酸盐时,可单独利用丙酸生长[20,21]。史密斯氏菌属(Smithella)具有氧化丙酸和丁酸的能力,通过延胡索酸激活了烷烃的起始代谢,并在此过程中与其他细菌共享氨基酸的合成代谢途径[20]。在GC11系统中,当系统有机负荷低于5.0 g VS/(L∙d) 时,互养棍状菌属、史密斯氏菌属和互营杆菌属的相对丰度分别为0.44% ~ 3.33%、0.99% ~ 4.35%和0.19% ~ 1.56%,而当有机负荷升高至5.5 g VS/(L·d) 时,其相对丰度分别显著降低至0.02%、0.09%和0.02%;而在GC31系统,当系统有机负荷低于4.5 g VS/(L∙d)时,这三种菌属的相对丰度分别为0.68% ~ 2.24%、1.33% ~ 2.75%和0.63% ~ 1.95%;继续升高有机负荷至5.0 ~ 5.5 g VS/(L∙d),其相对丰度分别降低至0.02% ~ 0.07%、0.08% ~ 0.10%和0.17% ~ 0.26%,其降幅达91.28% ~ 99.06%。结合贡献系统内丙酸和丁酸累积的菌属随有机负荷升高的变化趋势可见(图3),在有机负荷低于4.5 g VS/(L∙d)(GC31)或5.0 g VS/(L∙d)(GC11)时贡献系统内丙酸和丁酸累积的细菌菌属的相对丰度较低(平均值为0.53%),而参与丙酸/丁酸转化细菌菌属丰度较高(平均值为1.27%),可实现丙酸和丁酸的持续转化;当有机负荷在5.0 g VS/(L∙d)(GC31)或5.5 g VS/(L∙d)(GC11)时,贡献系统内丙酸和丁酸累积的细菌菌属的平均相对丰度升高至1.80%,而参与丙酸/丁酸转化细菌菌属平均丰度降低至0.13%,导致系统对丙酸/丁酸的转化效率降低,这与系统在高负荷阶段的丙酸和丁酸浓度增加有关[16]。
对于GC11系统,随着有机负荷从2.0 g VS/(L∙d)逐步增加至5.5 g VS/(L∙d),长蝇菌属(Levilinea)、Ornatilinea、排污管竿菌属(Cloacibacillus)和纤蝇菌属(Leptolinea)的相对丰度分别从7.37%降至1.20%、从1.35%降至0.14%、从1.17%降至0.21%和从0.45%降至0.13%;对于GC31系统,随着有机负荷升高,这4种菌属的相对丰度从3.68%降至1.30%、从0.65%降至0.29%、从1.81%降至0.58%和从1.99%降至0.04%。长蝇菌属、Ornatilinea、排污管竿菌属、纤蝇菌属均能参与蛋白质和氨基酸物质的转化。在GC11和GC31系统中,它们的相对丰度都呈现随着系统有机负荷的升高而降低的趋势。长蝇菌属可在pH值6.0 ~ 7.2时发酵糖类和氨基酸,其主要产物是H2、乙酸和乳酸;Ornatilinea可代谢多肽类和碳水化合物(包括微晶态纤维素),生长适宜pH值为7.5 ~ 8.0[22];而排污管竿菌属和纤蝇菌属都是蛋白和氨基酸降解菌[23,24]。在GC11系统中,当系统有机负荷低于4.5 g VS/(L∙d) 时,假单胞菌属(Pseudomonas)的相对丰度为0.06% ~ 0.93%,在5.0 ~ 5.5 g VS/(L∙d) 时相对丰度升高至3.68% ~ 5.78%;而在GC31系统中,该菌属的相对丰度则从系统有机负荷低于4.5 g VS/(L·d) 时的0.11% ~ 0.83%显著升至有机负荷为5.0 ~ 5.5 g VS/(L∙d) 时的1.07% ~ 3.54%,增幅可达29% ~ 3118%。当有机负荷从2.0 g VS/(L∙d) 升至5.5 g VS/(L∙d) 时,噬蛋白质菌属(Proteiniphilum)的相对丰度从0.00%升高至1.01% ~ 1.67%,且分别在有机负荷4.5 g VS/(L∙d) (GC11)和3.5 g VS/(L∙d) (GC31)时获得了最高的相对丰度,分别为3.36%和5.34%。假单胞菌属和噬蛋白质菌属与系统中氨氮积累有关。假单胞菌属可代谢葡萄糖和木糖,并且可分泌胞外蛋白酶强化不溶性蛋白的水解[25]。噬蛋白质菌属是专性厌氧蛋白质水解菌,可水解发酵酵母提取物、蛋白胨、丙酮酸、甘氨酸和精氨酸等,其主要的水解产物是乙酸和NH3[26,27]。可以看到,随着系统有机负荷升高,在系统失稳阶段,参与蛋白质和氨基酸物质转化的菌属相对丰度明显降低,而导致系统氨氮积累的菌属相对丰度显著升高,这与GC11和GC31系统在有机负荷为5.0 ~ 5.5 g VS/(L∙d) 时总氨氮(total ammonia nitrogen, TAN)浓度升高相对应[16]。
2.3 杂交狼尾草和牛粪混合发酵系统中古菌演替规律
在杂交狼尾草和牛粪混合发酵系统中,属水平上产甲烷菌在不同有机负荷和原料配比下的变化情况如图4所示。杂交狼尾草和牛粪混合发酵系统在有机负荷2.0 ~ 5.0 g VS/(L∙d)(GC11)或2.0 ~ 4.5 g VS/(L∙d)(GC31)时运行稳定,而在有机负荷为5.5 g VS/(L∙d) (GC11)或5.0 ~ 5.5 g VS/(L∙d) (GC31)时失稳[16]。与此对应地,在稳定运行时,GC11系统属水平上的优势产甲烷菌是甲烷丝菌属(Methanothrix),为乙酸营养型产甲烷菌[28],其相对丰度为40.16% ~ 57.34%,随着有机负荷的继续提高,系统失稳,优势产甲烷菌属转变为氢营养型产甲烷菌;而在GC31系统中,当有机负荷为2.0 g VS/(L∙d)时优势产甲烷菌属是甲烷蝇菌属(Methanolinea),为氢营养型产甲烷菌属[29],其相对丰度为37.27%,说明该系统在低有机负荷时主要利用氢产甲烷,随着有机负荷升高至2.5 ~ 4.5 g VS/(L∙d),甲烷丝菌属成为优势菌属,并且随着有机负荷升高,其相对丰度逐渐从55.11%降低至36.28%,说明在该有机负荷范围内系统主要利用乙酸产甲烷。在不同配比下系统稳定运行时甲烷八叠球菌属(Methanosarcina)的相对丰度均低于0.09%,而在系统失稳阶段,GC11系统有机负荷为5.5 g VS/(L∙d)以及GC31系统在5.0 ~ 5.5 g VS/(L∙d)时,其相对丰度显著升高至5.03%和4.56% ~ 9.40%。可见稳定运行阶段的杂交狼尾草和牛粪混合发酵系统以乙酸营养型产甲烷途径为主,氢营养型产甲烷途径为辅,系统中乙酸可持续转化,从而使乙酸维持较低浓度;当系统失稳时,乙酸营养型产甲烷菌属丰度降低,系统对乙酸利用性能降低,导致乙酸浓度增加,影响系统性能。
3 结论
(1)对于杂交狼尾草和牛粪混合发酵系统,在有机负荷低于4.5 g VS/(L∙d) (GC31)或5.0 g VS/(L∙d)(GC11)时贡献丙酸/丁酸累积的细菌菌属的相对丰度较低,而参与丙酸/丁酸转化细菌菌属丰度较高,可实现丙酸和丁酸的持续转化;同时贡献系统氨氮积累的假单胞菌属和噬蛋白质菌属丰度较低;而在产甲烷阶段,乙酸营养型产甲烷菌属是系统中的优势菌属,乙酸发酵是主要的产甲烷途径。
(2)当有机负荷升至5.0 g VS/(L∙d)或者 5.5 g VS/(L∙d) 时,贡献丙酸/丁酸累积的细菌菌属的相对丰度升高,而参与丙酸/丁酸转化细菌菌属丰度降低,系统对丙酸/丁酸的转化效率降低,与丙酸和丁酸浓度增加有关;此时与系统中氨氮浓度升高有关的假单胞菌属和噬蛋白质菌属丰度增加。在产甲烷阶段,乙酸营养型产甲烷菌属丰度降低,系统对乙酸利用性能降低,乙酸浓度增大。